
Post by Kate on Jun 24, 2009 23:29:15 GMT -5
This article is copyright Kate, 2009, and may not be reproduced in whole or in part without permission. This was originally written as part of my special project in fall 2008.
Genetics: Practical Applications as Related to the Equine Field
Here’s a problem; if you breed two bay horses together, what color will you get? If they were both genotypically EeAa, a simple pair of Punnett squares would give us the answer to what we might get.

According to Punnett square #1, you stand a 25% chance of having a horse dominant homozygous for E, 50% chance of having a horse who is heterozygous for E, and 25% chance of having a horse who is recessive homozygous for E.
According to Punnett square #2, we once again have a 25% chance of dominant homozygous, 50% chance of heterozygous, and 25% chance of recessive homozygous.
All of these combine to give us the following possible color combinations.
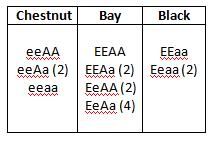
As a result, we can get any of three colors, but bay is the most likely possibility, with a 56.25% chance. Chestnut has a 25% chance, with black the least likely at 18.75%.
But what do all these terms mean? If you’ve taken biology, some might sound familiar, but for those who haven’t, let me take you through some of the basic terms.
- Genotype is the genetic makeup of a horse, or the actual genes present. A related term is phenotype, which is what the horse appears like. While a phenotype usually gives indication of what a horse’s genotype is, it’s not always accurate; for instance, the agouti gene is hidden in a chestnut horse, meaning that phenotypically the horse does not show agouti.
- Chromosomes determine the horse’s genotype. Every being has two sets of chromosomes, one from each parent. This causes two sets of alleles, or genes, that determine what colors are present. Each allele is represented by a letter (or two letters) that indicate certain color patterns.
(e)e-e(e)
(A)a-a(a)
(cr)Cr-cr(cr)
(g)g-G(g)
To-To
These chromosomes tell us that one parent was a palomino tobiano, while the other was a gray (chestnut based) tobiano. In turn, a knowledge of the effects of each allele the horse has (those not in parentheses) tells us that this horse is a gray tobiano with a palomino base.
- Homozygous is a term meaning that both alleles are the same – either dominant or recessive (XX or xx) (see below). Heterozygous is a term meaning that the alleles are different; one is dominant while the other is recessive (Xx).
- Alleles come in three different types. The first type, dominant, is the most prevalent. A dominant allele will always be apparent in the horse’s phenotype. As we’ll see, this includes alleles such as E, G, and D. Incomplete dominance means that one copy of the allele looks different from two copies, and both look different from no copies. Cr is perhaps the best-known of the incomplete dominant alleles. Recessive means that the color will only display phenotypically if there are no copies of the allele. Chestnut horses are the best example of this, as they must genetically be ee, or recessive homozygous.
These terms tend to occur a lot in the study of genetics, so if you’re not familiar with them, feel free to refer back to them; they’re going to be popping up quite a bit in the next few pages.
Now that we’ve covered some basic terms, we’re going to start discussing the real reason we’re here; what are these alleles, and how (and for that matter, why) do we learn about them? Let me give you a couple of practical examples. A good friend of mine breeds horses. She was telling me about a mare she has. Several generations back in this mare’s pedigree, one horse has both parents registered as chestnuts. The problem? The horse in question was registered as bay. Genetically, it is not possible for two chestnuts to produce a bay. This suggests one of two possibilities.
(a) One of the horses in question had their color misidentified or registered incorrectly.
(b) One of the parents listed (likely the sire) is not, in fact, the true parent.
So if you have a chestnut mare bred to a black stallion, and you wind up with a palomino foal, maybe you should check out who the other potential sires could be!
There are many varieties of alleles, but the first one I’m going to introduce you to is one of the building blocks of coat color – E, or eumelanin. Eumelanin is what causes black pigmentation in the coat. Horses come in essentially two colors – red-based colors such as chestnut, and black-based colors such as bay and black. Horses with a red-based coat are recessive homozygous for E [ee]. Horses with a black-based coat are either heterozygous [Ee] or dominant homozygous [EE] for E, thus giving them their black coloration. While most other alleles are excluded from written genotypes if they’re in a recessive homozygous form, E, as well as the next allele we’ll discuss, A, are always included in written genotypes, regardless of what form they take.
The A in question stands for agouti, which causes restriction of black. Agouti is a dominant gene, meaning that one copy (Aa) produces the same results as two copies (AA). Of the three basic coat colors (chestnut, bay, and black), A is only visible on bay, because it’s the reason that the black in the coat is restricted to only appearing on the mane, tail, and points of the horse. Black horses are always recessive homozygous (aa), and chestnut horses don’t have any black, so they can have any form of the allele. This ability to “hide” the allele is why breeders can occasionally be surprised when a black x chestnut breeding results in a bay foal. The bay foal is a sure indication that the chestnut parent is hiding at least one copy of the A allele.
Cream, or Cr, is the allele responsible for the colors of palomino, buckskin, and smoky black, on chestnut, bay, and black bases respectively (all alterations in color caused by an allele will be on those base colors in that order). The best way to describe the Cr allele is that it’s like adding creamer to coffee. It lightens the color, but it doesn’t remove it or override it completely. But what happens when you add a double dose of creamer to your coffee? It gives you an even lighter shade, which in horses, means that the dominant homozygous form [CrCr] produces the colors cremello, perlino, and smoky cream. Cr is an example of an incomplete dominant allele, where the physical traits differ depending on whether the allele is homozygous or heterozygous.
Similar in appearance to cream is champagne (Ch). Champagne dilutes the coat color as well, although since it is a dominant gene, one copy of the allele has the same effect as two. Champagne looks very similar to cream, but tends to give the coat a metallic sheen not seen with cream. Horses tend to be born with darker coats that will lighten with age. They’re also generally born with blue eyes that can change to green or hazel before becoming brown. To date, champagne has only been documented in Quarter Horses, Tennessee Walking Horses, American Saddlebreds, Missouri Fox Trotters, and Miniature Horses. Champagne produces the colors gold champagne, amber champagne, and classic champagne, on chestnut, bay, and black coats respectively.
Gray (G) is a dominant gene that is extremely common. In some breeds, such as Lipizzaners and Andalusians, horses are almost exclusively gray. Horses are not born gray, but rather turn gray as they age. Foals will be born the adult version of their color; for example, a bay foal will have black points instead of the usual light points. There are many varieties of gray, such as dappled, steel, iron, rose, and fleabitten. The exact reason as to why a horse is one pattern of gray instead of another is still unknown. Gray horses also tend to be prone to sunburn and melanomas as a result of sun exposure.
Roan is caused by the dominant Rn allele, and produces white hairs intermingled with the base coat. This trait is apparent when the foal coat sheds out. The horse will retain a solid head and lower legs, with the hairs at the knees forming an upward V of the solid base color.
The D allele produces the array of dun colors. Another dominant gene, D will produce a slight diluting effect on the coat, most notably on the bay and black bases, but it’s a unique gene in the fact that it causes what are known as primitive markings. These include darker areas on the face and neck known as shadowing, along with a black stripe down the back known as a dorsal stripe, an area of black over the withers known as shoulder striping, and dark points that turn into zebra-like striping on the legs. All of these may occur, or only a couple, but there will generally be some sort of primitive marking (most commonly the dorsal stripe) associated with the dun coloration. It produces red dun, bay (or yellow) dun, and grullo (or grulla in the case of a female). Bay dun can be quite similar to the buckskin produced by the Cr allele, although the primitive markings will make it clear that it is D and not Cr that is responsible for the coloration.
Dorsal stripes should not be confused with countershading. They look identical, but dorsal stripes are always associated with the dun gene, while countershading is not associated with any particular gene. Countershading in conjunction with buckskin could be potentially mistaken for a yellow dun, but the parentage of the foal should make it clear if this is the case – otherwise, genetic testing can be used.
Silver dapple (the Z allele) causes the retention of black pigment. This is widespread in breeds such as the Rocky Mountain Horse and Kentucky Mountain Horse, and has also been observed recently in the Morgan, Quarter Horse, and Arabian breeds. It’s also known to exist in Icelandic Horses, American Shetland Ponies, and Miniature Horses. Because it is limited to the black pigment only, it’s hidden on a red-based horse. Bay horses will have coats which are distinctly red while black horses will have coats that are closer to a chocolate brown. It will cause a flaxen mane and tail with both colors. Z has been linked to eye problems, including moon blindness.
The leopard allele (Lp) is a dominant gene which causes expression of spotted patterns, most commonly associated with Appaloosas but also seen in breeds such as the Miniature Horse and Knabstrupper. It is unique due to the fact that it can, apparently at random, produce five different pattern types. Minimal expression will include mottled skin, especially on the muzzle, striped hooves, and white sclera in the eyes. Maximum expression can produce a horse with only a few spots of color.
Tobiano (To) is a pinto gene, causing horses to have large splashes of white that cross their topline. In addition, horses will commonly have four white socks, and a solid head. The white will tend to stretch vertically from the top of the barrel to the bottom. There may also be small spots located in the white area, known as ink spots, paw prints, or cat tracks. Related to tobiano are the overo genes. Frame overo (Fr or O) will cause white that is located on the sides of the barrel and the neck. It will not cross the topline. Frame overo horses will generally have a lot of white on their heads and minimal white on their legs, the opposite of tobiano. Splash overo (Spl) horses will have a lot of white on their bellies and legs, extending up their sides. They will also generally have white faces. Spl is the only one of these three that differs based on whether it is homozygous or heterozygous; horses who are dominant homozygous for Spl will have more white than horses who are heterozygous. However, all these genes are dominant and will be expressed if present.
Sabino (Sb) is another pinto gene, but it’s somewhat of a mystery. It is expressed via abundant white on the legs, spots on the belly or body that tend to be flecked or look somewhat roan, chin spots, or white on the face that surrounds the eyes. It can be expressed in a minimal form which means horses can have a small spot on their belly or body, or long stockings on their legs. It can also be expressed in a maximal form which turns the whole horse white. Minimally-expressed horses have been known to produce maximally-expressed horses. This also appears to be polygenic in nature (having more than one gene that causes it), as the gene identified for causing sabino in most breeds has also been identified as not being the cause in Arabians and Clydesdales, two breeds where sabino horses are common.
White (W) is a very unique gene. Relatively rare, it can be easily confused with maximum sabino, but since W is a dominant allele, one parent must be white in order for the horse to be white. It is not a proper albino, since the horse will have blue eyes (true albino, which include pink eyes, does not exist in horses). W is lethal in dominant homozygous form (WW) and will mask any other color genes present, requiring genetic testing to determine what is present.
There are many other color genes present in horses, such as rabicano (Rb), pangare (P), non-fading black (E+), pearl (Prl), flaxen (F), and sooty (Sty). The full range of equine coat color has still not been determined, as shown by the polygenic theory with Sb, the recent discovery of Prl, and the fact that E+ is still somewhat hypothetical. Equine color coat genetics is a fascinating and complex subject with a very practical use, and I strongly encourage all those involved with horses, in whatever way, to study it.
Works Cited
"Chart of Color Alleles." Model Horse Reference. 31 Aug. 2008 <http://mhref.com/color/genetics/loci.html>.
Equine Color. 31 Aug. 2008 <http://equinecolor.com>.
"Horse Coat Color." Veterinary Genetics Laboratory. University of California - Davis. 14 Dec. 2008 <http://www.vgl.ucdavis.edu/services/coatcolorhorse.php>.
Sponenberg, D. Phillip. Equine Color Genetics. 2nd ed. Grand Rapids: Blackwell Limited, 2003.
Genetics: Practical Applications as Related to the Equine Field
Here’s a problem; if you breed two bay horses together, what color will you get? If they were both genotypically EeAa, a simple pair of Punnett squares would give us the answer to what we might get.
According to Punnett square #1, you stand a 25% chance of having a horse dominant homozygous for E, 50% chance of having a horse who is heterozygous for E, and 25% chance of having a horse who is recessive homozygous for E.
According to Punnett square #2, we once again have a 25% chance of dominant homozygous, 50% chance of heterozygous, and 25% chance of recessive homozygous.
All of these combine to give us the following possible color combinations.
As a result, we can get any of three colors, but bay is the most likely possibility, with a 56.25% chance. Chestnut has a 25% chance, with black the least likely at 18.75%.
But what do all these terms mean? If you’ve taken biology, some might sound familiar, but for those who haven’t, let me take you through some of the basic terms.
- Genotype is the genetic makeup of a horse, or the actual genes present. A related term is phenotype, which is what the horse appears like. While a phenotype usually gives indication of what a horse’s genotype is, it’s not always accurate; for instance, the agouti gene is hidden in a chestnut horse, meaning that phenotypically the horse does not show agouti.
- Chromosomes determine the horse’s genotype. Every being has two sets of chromosomes, one from each parent. This causes two sets of alleles, or genes, that determine what colors are present. Each allele is represented by a letter (or two letters) that indicate certain color patterns.
(e)e-e(e)
(A)a-a(a)
(cr)Cr-cr(cr)
(g)g-G(g)
To-To
These chromosomes tell us that one parent was a palomino tobiano, while the other was a gray (chestnut based) tobiano. In turn, a knowledge of the effects of each allele the horse has (those not in parentheses) tells us that this horse is a gray tobiano with a palomino base.
- Homozygous is a term meaning that both alleles are the same – either dominant or recessive (XX or xx) (see below). Heterozygous is a term meaning that the alleles are different; one is dominant while the other is recessive (Xx).
- Alleles come in three different types. The first type, dominant, is the most prevalent. A dominant allele will always be apparent in the horse’s phenotype. As we’ll see, this includes alleles such as E, G, and D. Incomplete dominance means that one copy of the allele looks different from two copies, and both look different from no copies. Cr is perhaps the best-known of the incomplete dominant alleles. Recessive means that the color will only display phenotypically if there are no copies of the allele. Chestnut horses are the best example of this, as they must genetically be ee, or recessive homozygous.
These terms tend to occur a lot in the study of genetics, so if you’re not familiar with them, feel free to refer back to them; they’re going to be popping up quite a bit in the next few pages.
Now that we’ve covered some basic terms, we’re going to start discussing the real reason we’re here; what are these alleles, and how (and for that matter, why) do we learn about them? Let me give you a couple of practical examples. A good friend of mine breeds horses. She was telling me about a mare she has. Several generations back in this mare’s pedigree, one horse has both parents registered as chestnuts. The problem? The horse in question was registered as bay. Genetically, it is not possible for two chestnuts to produce a bay. This suggests one of two possibilities.
(a) One of the horses in question had their color misidentified or registered incorrectly.
(b) One of the parents listed (likely the sire) is not, in fact, the true parent.
So if you have a chestnut mare bred to a black stallion, and you wind up with a palomino foal, maybe you should check out who the other potential sires could be!
There are many varieties of alleles, but the first one I’m going to introduce you to is one of the building blocks of coat color – E, or eumelanin. Eumelanin is what causes black pigmentation in the coat. Horses come in essentially two colors – red-based colors such as chestnut, and black-based colors such as bay and black. Horses with a red-based coat are recessive homozygous for E [ee]. Horses with a black-based coat are either heterozygous [Ee] or dominant homozygous [EE] for E, thus giving them their black coloration. While most other alleles are excluded from written genotypes if they’re in a recessive homozygous form, E, as well as the next allele we’ll discuss, A, are always included in written genotypes, regardless of what form they take.
The A in question stands for agouti, which causes restriction of black. Agouti is a dominant gene, meaning that one copy (Aa) produces the same results as two copies (AA). Of the three basic coat colors (chestnut, bay, and black), A is only visible on bay, because it’s the reason that the black in the coat is restricted to only appearing on the mane, tail, and points of the horse. Black horses are always recessive homozygous (aa), and chestnut horses don’t have any black, so they can have any form of the allele. This ability to “hide” the allele is why breeders can occasionally be surprised when a black x chestnut breeding results in a bay foal. The bay foal is a sure indication that the chestnut parent is hiding at least one copy of the A allele.
Cream, or Cr, is the allele responsible for the colors of palomino, buckskin, and smoky black, on chestnut, bay, and black bases respectively (all alterations in color caused by an allele will be on those base colors in that order). The best way to describe the Cr allele is that it’s like adding creamer to coffee. It lightens the color, but it doesn’t remove it or override it completely. But what happens when you add a double dose of creamer to your coffee? It gives you an even lighter shade, which in horses, means that the dominant homozygous form [CrCr] produces the colors cremello, perlino, and smoky cream. Cr is an example of an incomplete dominant allele, where the physical traits differ depending on whether the allele is homozygous or heterozygous.
Similar in appearance to cream is champagne (Ch). Champagne dilutes the coat color as well, although since it is a dominant gene, one copy of the allele has the same effect as two. Champagne looks very similar to cream, but tends to give the coat a metallic sheen not seen with cream. Horses tend to be born with darker coats that will lighten with age. They’re also generally born with blue eyes that can change to green or hazel before becoming brown. To date, champagne has only been documented in Quarter Horses, Tennessee Walking Horses, American Saddlebreds, Missouri Fox Trotters, and Miniature Horses. Champagne produces the colors gold champagne, amber champagne, and classic champagne, on chestnut, bay, and black coats respectively.
Gray (G) is a dominant gene that is extremely common. In some breeds, such as Lipizzaners and Andalusians, horses are almost exclusively gray. Horses are not born gray, but rather turn gray as they age. Foals will be born the adult version of their color; for example, a bay foal will have black points instead of the usual light points. There are many varieties of gray, such as dappled, steel, iron, rose, and fleabitten. The exact reason as to why a horse is one pattern of gray instead of another is still unknown. Gray horses also tend to be prone to sunburn and melanomas as a result of sun exposure.
Roan is caused by the dominant Rn allele, and produces white hairs intermingled with the base coat. This trait is apparent when the foal coat sheds out. The horse will retain a solid head and lower legs, with the hairs at the knees forming an upward V of the solid base color.
The D allele produces the array of dun colors. Another dominant gene, D will produce a slight diluting effect on the coat, most notably on the bay and black bases, but it’s a unique gene in the fact that it causes what are known as primitive markings. These include darker areas on the face and neck known as shadowing, along with a black stripe down the back known as a dorsal stripe, an area of black over the withers known as shoulder striping, and dark points that turn into zebra-like striping on the legs. All of these may occur, or only a couple, but there will generally be some sort of primitive marking (most commonly the dorsal stripe) associated with the dun coloration. It produces red dun, bay (or yellow) dun, and grullo (or grulla in the case of a female). Bay dun can be quite similar to the buckskin produced by the Cr allele, although the primitive markings will make it clear that it is D and not Cr that is responsible for the coloration.
Dorsal stripes should not be confused with countershading. They look identical, but dorsal stripes are always associated with the dun gene, while countershading is not associated with any particular gene. Countershading in conjunction with buckskin could be potentially mistaken for a yellow dun, but the parentage of the foal should make it clear if this is the case – otherwise, genetic testing can be used.
Silver dapple (the Z allele) causes the retention of black pigment. This is widespread in breeds such as the Rocky Mountain Horse and Kentucky Mountain Horse, and has also been observed recently in the Morgan, Quarter Horse, and Arabian breeds. It’s also known to exist in Icelandic Horses, American Shetland Ponies, and Miniature Horses. Because it is limited to the black pigment only, it’s hidden on a red-based horse. Bay horses will have coats which are distinctly red while black horses will have coats that are closer to a chocolate brown. It will cause a flaxen mane and tail with both colors. Z has been linked to eye problems, including moon blindness.
The leopard allele (Lp) is a dominant gene which causes expression of spotted patterns, most commonly associated with Appaloosas but also seen in breeds such as the Miniature Horse and Knabstrupper. It is unique due to the fact that it can, apparently at random, produce five different pattern types. Minimal expression will include mottled skin, especially on the muzzle, striped hooves, and white sclera in the eyes. Maximum expression can produce a horse with only a few spots of color.
Tobiano (To) is a pinto gene, causing horses to have large splashes of white that cross their topline. In addition, horses will commonly have four white socks, and a solid head. The white will tend to stretch vertically from the top of the barrel to the bottom. There may also be small spots located in the white area, known as ink spots, paw prints, or cat tracks. Related to tobiano are the overo genes. Frame overo (Fr or O) will cause white that is located on the sides of the barrel and the neck. It will not cross the topline. Frame overo horses will generally have a lot of white on their heads and minimal white on their legs, the opposite of tobiano. Splash overo (Spl) horses will have a lot of white on their bellies and legs, extending up their sides. They will also generally have white faces. Spl is the only one of these three that differs based on whether it is homozygous or heterozygous; horses who are dominant homozygous for Spl will have more white than horses who are heterozygous. However, all these genes are dominant and will be expressed if present.
Sabino (Sb) is another pinto gene, but it’s somewhat of a mystery. It is expressed via abundant white on the legs, spots on the belly or body that tend to be flecked or look somewhat roan, chin spots, or white on the face that surrounds the eyes. It can be expressed in a minimal form which means horses can have a small spot on their belly or body, or long stockings on their legs. It can also be expressed in a maximal form which turns the whole horse white. Minimally-expressed horses have been known to produce maximally-expressed horses. This also appears to be polygenic in nature (having more than one gene that causes it), as the gene identified for causing sabino in most breeds has also been identified as not being the cause in Arabians and Clydesdales, two breeds where sabino horses are common.
White (W) is a very unique gene. Relatively rare, it can be easily confused with maximum sabino, but since W is a dominant allele, one parent must be white in order for the horse to be white. It is not a proper albino, since the horse will have blue eyes (true albino, which include pink eyes, does not exist in horses). W is lethal in dominant homozygous form (WW) and will mask any other color genes present, requiring genetic testing to determine what is present.
There are many other color genes present in horses, such as rabicano (Rb), pangare (P), non-fading black (E+), pearl (Prl), flaxen (F), and sooty (Sty). The full range of equine coat color has still not been determined, as shown by the polygenic theory with Sb, the recent discovery of Prl, and the fact that E+ is still somewhat hypothetical. Equine color coat genetics is a fascinating and complex subject with a very practical use, and I strongly encourage all those involved with horses, in whatever way, to study it.
Works Cited
"Chart of Color Alleles." Model Horse Reference. 31 Aug. 2008 <http://mhref.com/color/genetics/loci.html>.
Equine Color. 31 Aug. 2008 <http://equinecolor.com>.
"Horse Coat Color." Veterinary Genetics Laboratory. University of California - Davis. 14 Dec. 2008 <http://www.vgl.ucdavis.edu/services/coatcolorhorse.php>.
Sponenberg, D. Phillip. Equine Color Genetics. 2nd ed. Grand Rapids: Blackwell Limited, 2003.





